Il trasportatore universale di energia metabolica nei sistemi biologici è l’ATP (Adenosina trifosfato). In realtà non esiste solo l’ATP come molecola energetica; ci sono molecole che contengono un livello di energia anche più elevato rispetto a quello dell’ATP. La differenza tra queste molecole e l’ATP è che le cellule, durante il corso dell’evoluzione, hanno premiato l’ATP come trasportatore universale di energia anche se i sistemi biologici avrebbero potuto scegliere un altro tipo di molecola.
Le scelte evolutive in molti casi sono casuali e probabilmente è stata casuale la scelta dell’ATP come trasportatore universale di energia.
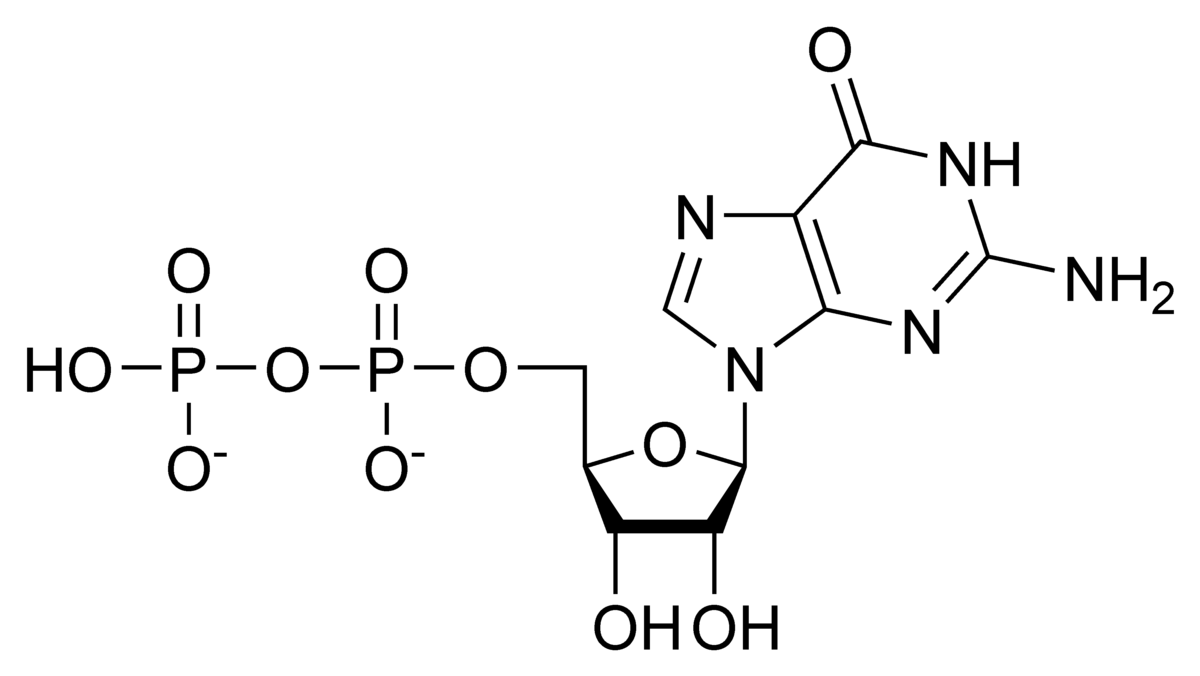
Un qualsiasi nucleotide è formato da una base azotata, legata con legame N-β-glicosidico con la porzione centrale del nucleotide che è uno zucchero pentosio, sul carbonio numero 1 dello zucchero. Sul carbonio numero 5, invece, si lega la porzione restante del nucleotide, ossia i gruppi fosfato.
I legami fosfo-anidridici tengono uniti il secondo e il terzo gruppo fosfato che si chiamano rispettivamente β e γ. Oltre all’ATP può essere utilizzato come trasportatore il GTP che è isoenergetico in quanto l’energia non deriva dalla differente base azotata ma dai gruppi fosforici, che sono uguali. Quando si rompe un legame fosfo-anidridico vengono liberati circa 30,5 kJ/mol, corrispondenti a circa 7,5 kcal/mol (1 kcal = 4,186 kJ).

In una molecola di ATP o di GTP o in un qualsiasi nucleoside trifosfato abbiamo a disposizione due legami fosfo-anidridici, quindi 30,5 kJ/mol va moltiplicato per due. Il legame tra il primo gruppo fosfato (α) e lo zucchero è fosfodiesterico: si forma tra un carbonio alcolico (carbonio 5 del ribosio nell’ATP) e il gruppo fosforico (paragonabile, reattivamente, a un gruppo carbossilico). Questa reazione chimica è caratterizzata da un contenuto di energia inferiore rispetto al legame fosfo-anidridico.
Altre molecole trasportatrici di energia (contenitori temporanei di energia che servono poi per la sintesi di ATP) sono le forme ridotte dei coenzimi NAD+ e FAD+ che, quando si riducono, diventano NADH + H+ e FADH2.
Tutte le reazioni che avvengono nelle cellule hanno degli scopi ben precisi, nulla avviene a caso. Soltanto molto raramente si possono percorrere all’interno della cellula i cosiddetti cicli futili. Per esempio degradare una molecola per riformarla subito dopo può servire soltanto per generare del calore all’interno nella cellula.

Le reazioni redox sono caratterizzate da un flusso di elettroni, che si traduce in flusso di energia; oltre al flusso di elettroni c’è uno spostamento di protoni. In biochimica tutte le reazioni redox prevedono uno spostamento di elettroni accompagnato da uno spostamento di protoni. Poiché un elettrone e un protone costituiscono l’atomo di idrogeno, durante queste fasi redox del metabolismo c’è uno spostamento di atomi di idrogeno. Tanto è vero che moltissimi enzimi che catalizzano le reazioni redox hanno l’appellativo di deidrogenasi, cioè rimuovono uno o più atomi di idrogeno da un substrato spostandolo/i su un altro substrato per formare il prodotto.
Quindi i coenzimi ridotti NADH e FADH2 rappresentano dei trasportatori temporanei di energia. Sono capaci di sintetizzare varie molecole di ATP per la modalità con cui trasferiscono energia alla catena di trasporto degli elettroni.
Dal loro contenuto energetico, per le leggi della termodinamica secondo le quali non tutta l’energia che è presente su una molecola o che comunque è a disposizione può essere trasformata in lavoro, possiamo ottenere 2,5 molecole di ATP dal NADH e un po’ meno dal FADH2 (dipende dal punto di ingresso nella catena respiratoria).

{kind=link}
Nella cellula esistono dei substrati come il fosfoenolpiruvato ad elevato contenuto energetico, anche chiamati substrati ad elevato potenziale di trasferimento del gruppo fosforico. Anche se sono meglio descritti come “ad elevato contenuto di energia” perché quello che viene trasferito non è tanto il gruppo fosforico quanto l’energia che serve per formare il legame fosfo-anidridico. Il fosfoenolpiruvato è un intermedio della via glicolitica, così come l’1-3-bisfosfoglicerato, ed è ad alto contenuto energetico. Il contenuto energetico di queste due molecole (circa 60 kJ/mol) è molto maggiore dell’ATP perché questa energia che viene liberata non può essere utilizzata al 100% secondo i principi della termodinamica. Un altro esempio classico di substrato ad alto contenuto energetico è il succinil-coenzima A che troviamo nel ciclo di Krebs e la cui energia risiede nel legame tioestere che si forma tra uno dei gruppi carbossilici del succinato e il gruppo tiolico del coenzima A. La fosfocreatina, che si trova a livello muscolare, è un magazzino di energia per l’apparato muscolare.