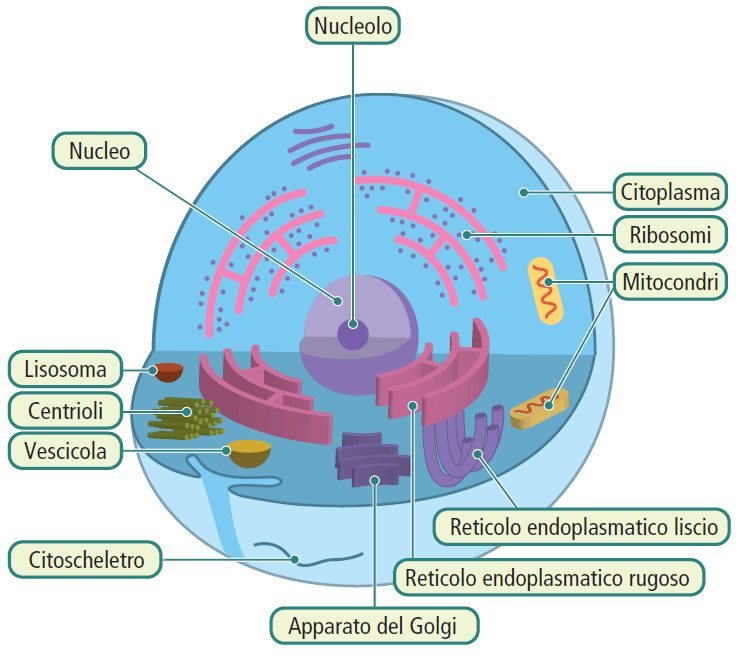
Il termine “eucariote” deriva dal greco e significa “vero nucleo”. Le cellule eucariotiche si distinguono per la presenza di un nucleo ben definito, circondato da un involucro membranoso chiamato involucro nucleare. A differenza delle cellule procariotiche, le cellule eucariotiche presentano una compartimentazione interna, con numerosi compartimenti intracellulari distinti da membrane. Gli eucarioti possiedono anche molteplici molecole di DNA lineare, che si compattano formando strutture chiamate cromosomi grazie all’interazione con proteine chiamate istoni.
Fonte: Manuale di preclinica.
All’interno del nucleo si trova il nucleolo, dove viene sintetizzato l’RNA ribosomiale. Nel citoplasma, esistono “autostrade” costituite da proteine che formano il citoscheletro, consentendo il movimento degli organuli citoplasmatici.
Gli organismi formati da cellule eucariotiche possono essere unicellulari (alcuni protisti) o pluricellulari (funghi, piante, animali).
La cellula eucariotica è composta da diverse componenti:
- Membrana plasmatica;
- Nucleo;
- Citoplasma;
- Organuli cellulari:
- Mitocondri;
- Apparato di Golgi;
- Lisosomi;
- Reticolo endoplasmatico;
- Vacuoli;
- Cloroplasti (presenti solo nelle cellule vegetali);
- Centrioli;
- Citoscheletro.

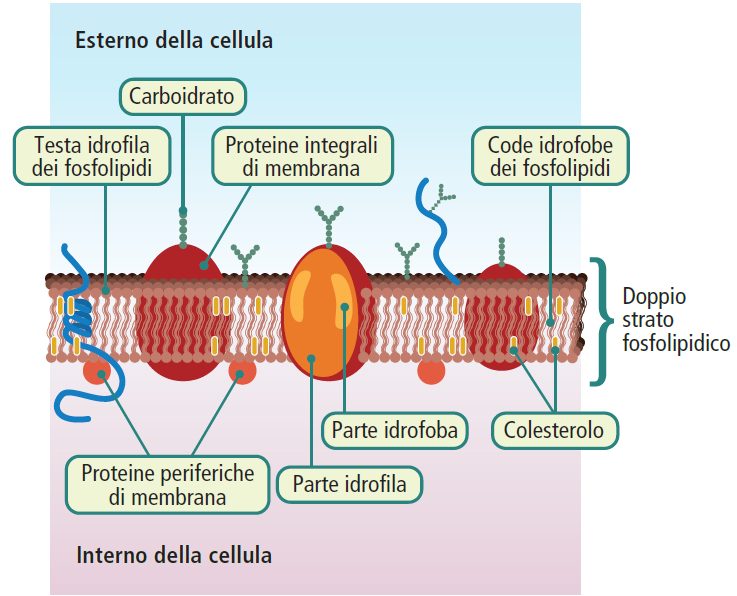
Membrana plasmatica
La membrana plasmatica separa l’ambiente interno da quello esterno, mantenendo un ambiente adatto allo svolgimento delle reazioni chimiche vitali.
Fonte: Manuale di preclinica.
Tutte le membrane biologiche, inclusa la membrana plasmatica, sono composte da un doppio strato di fosfolipidi, proteine e una piccola quantità di carboidrati.
I fosfolipidi, i lipidi predominanti, sono costituiti da una parte idrofoba a forma di “coda” con due catene di acidi grassi e una parte idrofila a forma di “testa” contenente un gruppo fosfato. Questa struttura anfipatica permette ai fosfolipidi di organizzarsi formando un doppio strato in acqua. Altri lipidi presenti nelle membrane delle cellule eucariotiche includono il colesterolo (20%), che regola la fluidità della membrana, e i glicolipidi (5%).
Le proteine presenti nella membrana possono essere suddivise in due categorie: periferiche e integrali. Le proteine periferiche sono debolmente associate al doppio strato fosfolipidico, mentre le proteine integrali attraversano completamente o parzialmente la membrana.
Le proteine svolgono diverse funzioni, come funzioni strutturali, di trasporto (le proteine integrali agiscono come canali per il passaggio di specifici sostanze), di riconoscimento e di comunicazione (le proteine recettoriali sono spesso glicoproteine che sporgono all’esterno della membrana plasmatica). In generale, maggiore è l’attività metabolica della membrana cellulare, maggiore sarà il contenuto proteico.
I carboidrati partecipano al processo di glicosilazione di proteine (glicoproteine) e lipidi (glicolipidi) e svolgono diversi ruoli, come la lubrificazione, l’adesione, la funzione recettoriale e la partecipazione alla formazione della matrice extracellulare (ECM, extracellular matrix).
Il modello attualmente accettato per le membrane biologiche è il modello del “mosaico fluido“, che descrive una struttura flessibile in cui lipidi e proteine possono spostarsi liberamente all’interno del piano della membrana.

Citoplasma
Il termine citoplasma indica l’insieme dei componenti contenuti all’interno della membrana cellulare di una cellula, che si trovano sia nelle cellule eucariotiche che in quelle procariotiche. Negi eucarioti, il citoplasma è composto da diverse parti: il citosol, il citoscheletro e gli organuli cellulari, che costituiscono la parte corpuscolata.
Sinonimi di citoplasma includono il termine sarcoplasma, utilizzato per indicare il citoplasma delle cellule muscolari, e assoplasma, che si riferisce al citoplasma dei neuroni.
Il citosol è la matrice fluida. La consistenza gelatinosa è attribuita alla presenza di una grande quantità di proteine disciolte. Oltre alle proteine, sono presenti anche piccole molecole come amminoacidi, zuccheri semplici e nucleotidi, che sono necessari per il corretto funzionamento degli organuli cellulari. Nel citosol si trovano anche metaboliti, che sono i prodotti intermedi della sintesi e della degradazione delle macromolecole, ioni inorganici che possono essere legati a proteine specifiche per il loro trasporto, e molecole ad alta energia come l’ATP.
Il citosol rappresenta circa il 50% del volume totale della cellula ed è composto per il 70% da acqua. Mantiene un valore di pH vicino alla neutralità, intorno a 7,0. Oltre a svolgere un ruolo importante nella sintesi proteica, il citosol ospita numerose vie metaboliche.
Fonte: Manuale di preclinica.
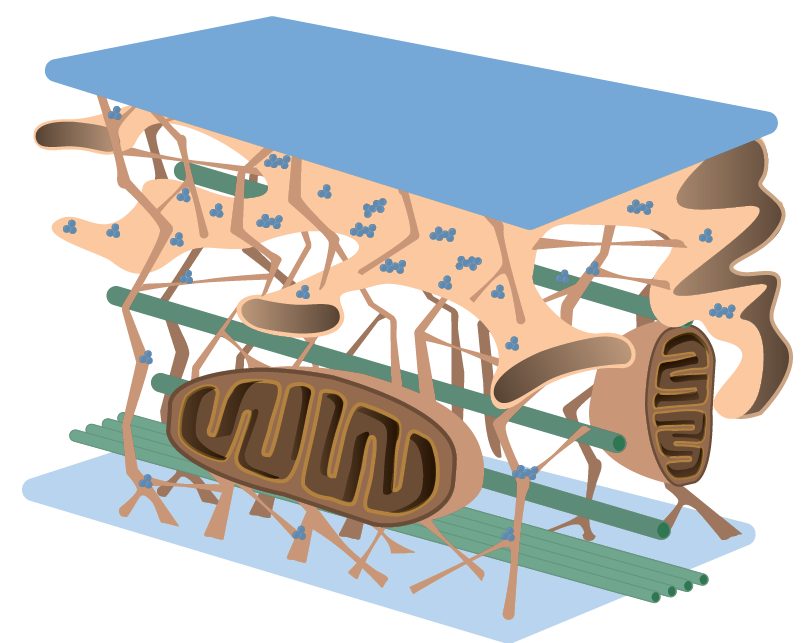
Il citoscheletro è un sistema complesso di strutture che si trovano sia all’interno che all’esterno del citoplasma delle cellule eucariotiche, e insieme formano una rete tridimensionale che funge da impalcatura. Tuttavia, il citoscheletro non è solo una struttura statica, ma è altamente dinamico e svolge diverse funzioni vitali all’interno della cellula. È responsabile della forma, del movimento e delle connessioni cellulari, partecipa al movimento degli organuli e alla divisione cellulare, e fornisce resistenza alla trazione.
Il citoscheletro è composto principalmente da tre tipi di filamenti proteici:
- I microfilamenti sono i filamenti più sottili, con uno spessore di circa 6-7 nm, formati da polimeri elicoidali flessibili di actina, noti anche come filamenti di actina. L’actina è una proteina globulare che si polimerizza attraverso l’aggiunta di unità monomeriche di actina legate all’ATP. I microfilamenti mostrano una polarità strutturale, con un’estremità positiva chiamata “barbed end“, dove avviene principalmente l’aggiunta di monomeri di G-actina per l’allungamento del filamento, e un’estremità negativa chiamata “pointed end“, dove prevalgono i processi di depolarizzazione dell’actina. L’accrescimento del filamento è un processo attivo che richiede l’idrolisi dell’ATP in ADP.
I microfilamenti sono coinvolti nella formazione strutturale dei microvilli, nei movimenti cellulari, nell’endocitosi, nella citodieresi e nella contrazione muscolare. - I filamenti intermedi devono il loro nome allo spessore, di circa 10 nm, intermedio tra i microfilamenti di actina e i microtubuli. Le proteine fibrose che costituiscono i filamenti intermedi variano a seconda del tipo di cellula, rendendo questa categoria di filamenti eterogenea. Ad esempio, nell’epitelio si trovano cheratine, nel tessuto muscolare o connettivo si trovano vimentine e nei neuroni si trovano neurofilamenti.
I filamenti intermedi sono caratterizzati da una grande resistenza alla trazione, che consente alla cellula di sopportare lo stress meccanico. A differenza degli altri filamenti del citoscheletro, non mostrano polarità e sono più stabili. I filamenti intermedi si polimerizzano attraverso l’aggregazione di monomeri che formano dimeri, che a loro volta si uniscono lateralmente per formare tetrameri. Infine, i tetrameri si aggregano per formare un filamento costituito da 32 monomeri, simile a un filo di corda. I tetrameri si associano lateralmente in strutture piane che successivamente si ripiegano in strutture cave.
Un esempio di filamenti intermedi presenti in tutte le cellule sono le lamine, che costituiscono la lamina nucleare, una rete di filamenti intermedi che sostiene il nucleo. Durante il processo di divisione cellulare, le proteine che costituiscono i filamenti intermedi della lamina nucleare vengono fosforilate dalle proteine chinasi, rendendoli instabili e favorendo la loro depolimerizzazione.
I filamenti intermedi possono interagire con diverse proteine, tra cui la desmoplachina, che li collega ai desmosomi e agli emidesmosomi, la plectrina, che li collega ai microtubuli, e l’anchirina, che li collega ai microfilamenti. - I microtubuli sono strutture proteiche cilindriche e cave composte da filamenti di tubulina. Essi si originano da un centro organizzatore dei microtubuli chiamato MTOC (centrosoma), localizzato vicino al nucleo. Il centrosoma è costituito dai due centrioli disposti perpendicolarmente, dal materiale pericentriolare e dai microtubuli astrali (aster). Ogni centriolo è composto da 9 triplette di microtubuli, da cui si originano nuovi microtubuli. I microtubuli formano una struttura rigida con un diametro di circa 25 nm. Sono composti da eterodimeri di tubulina α e tubulina β. Oltre a queste due isoforme, esiste anche la tubulina γ, che svolge un ruolo importante nel processo di nucleazione dei dimeri di α- e β-tubulina, oltre a formare un anello collegato ai microtubuli in fase di allungamento. Il centrosoma contiene anelli di tubulina γ che agiscono come siti di innesco (nucleazione) per la crescita dei microtubuli. I microtubuli si allungano e si accorciano in un equilibrio dinamico dipendente dall’GTP. Solo la tubulina β può idrolizzare l’GTP in GDP.
I microtubuli presentano una polarità: un’estremità (+) in corrispondenza della subunità β e un’estremità (-) in corrispondenza della subunità α. Alle estremità (+), l’aggiunta dei dimeri è quantitativamente superiore alla loro rimozione, determinando un allungamento del polimero. La situazione è opposta alle estremità (-).
Oltre al ruolo strutturale, i microtubuli costituiscono il fuso mitotico necessario per il movimento dei cromosomi durante la mitosi e la meiosi. Essi formano anche ciglia e flagelli, che consentono il movimento di alcune cellule, il traffico vescicolare e l’ancoraggio degli organelli al citoplasma. Le ciglia, presenti soprattutto nelle vie respiratorie, sono più corte e numerose rispetto ai flagelli (ad esempio, nello spermatozoo è presente un unico grande flagello). La struttura di ciglia e flagelli è simile, con microtubuli disposti secondo un modello 9+2 (nove coppie di microtubuli disposte a cerchio attorno a una coppia di microtubuli centrali singoli). Ciglia e flagelli sono ancorati alla cellula tramite un corpo basale con una struttura 9×3 (cioè nove triplette di microtubuli disposte a cerchio, simili ai centrioli).
Alcuni composti influenzano il comportamento dei microtubuli:- La colchicina, un alcaloide derivato dalla pianta Colchicum autumnale, ha un’azione chemioterapica. Si lega a una singola molecola di tubulina ma non alla tubulina polimerizzata. Inibisce lo scambio di subunità e causa la disgregazione del fuso mitotico, bloccando la mitosi.
- Il taxolo, estratto dalla corteccia del Taxus brevifolia, ha anche un’azione chemioterapica ma con un meccanismo opposto alla colchicina. Si lega ai microtubuli e li stabilizza. Le cellule si arrestano in mitosi, evidenziando la necessità che i microtubuli si depolimerizzino anche per rendere funzionale il fuso.

Nucleo
Il nucleo cellulare è considerato l’organello più fondamentale all’interno della cellula. Presenta una forma sferica o ovoidale con un diametro medio di circa 5 μm. È costituito da due membrane concentriche che separano il suo contenuto dal citoplasma circostante. Queste membrane si fondono a intervalli regolari formando dei pori nucleari che consentono il passaggio selettivo di materiali tra il nucleo e il citoplasma.
Il nucleo svolge una serie di funzioni cruciali all’interno della cellula. Uno dei suoi compiti principali è quello di contenere il DNA, che si associa a proteine fondamentali chiamate istoni e altre proteine acide per formare la cromatina. Durante la divisione cellulare, la cromatina si condensa formando strutture ben definite chiamate cromosomi.
Oltre al DNA e alla cromatina, il nucleo ospita anche una regione chiamata nucleolo. Il nucleolo svolge un ruolo essenziale nella sintesi dell’RNA ribosomiale e nell’assemblaggio dei ribosomi. Nonostante non sia considerato un vero e proprio organulo interno al nucleo, il nucleolo rappresenta una regione densa di materiale genetico e proteico che può essere osservata sia al microscopio ottico che al microscopio elettronico.
Ribosomi
I ribosomi nelle cellule eucariotiche, ad eccezione di quelli contenuti nei mitocondri e nei cloroplasti, sono più grandi rispetto a quelli presenti nelle cellule procariotiche. Hanno una massa molecolare di circa 4.000 kDa, un diametro di 23 nm e un coefficiente di sedimentazione di 80S. Sono composti da due subunità, una maggiore (60S) e una minore (40S):
- La subunità maggiore è costituita da tre molecole di RNA ribosomiale (rRNA): una di 28S, una di 5,8S e una più piccola di 5S.
- La subunità minore è composta da una singola catena di rRNA di 18S.
Complessivamente, le due subunità sono composte da oltre 80 proteine. Le molecole individuali di rRNA (ad eccezione del 5S) vengono sintetizzate nei nucleoli come RNA 45S. Il DNA presente nel nucleolo viene trascritto dalla RNA polimerasi I da diversi punti del DNA, formando strutture chiamate “cistroni“, che assomigliano a un “albero di Natale” in cui il tronco rappresenta il DNA e i rami rappresentano le numerose catene di rRNA trascritte contemporaneamente.
I ribosomi liberi si trovano nel citoplasma cellulare o fanno parte dei mitocondri, a differenza di quelli associati alla membrana esterna del reticolo endoplasmatico rugoso, che sintetizzano e rilasciano proteine all’interno delle membrane di queste strutture. Successivamente, le proteine vengono indirizzate verso la loro destinazione finale, che può essere sia all’interno che all’esterno della cellula (nel caso di secrezione cellulare attraverso l’esocitosi).
Le due subunità del ribosoma si uniscono e collaborano per tradurre l’RNA messaggero in una catena polipeptidica durante la sintesi proteica. Sembra che le molecole di RNA ribosomiale siano la parte più importante dei ribosomi in termini di traduzione, anche se il processo avviene lentamente. L’azione delle componenti proteiche sembra potenziare e accelerare l’attività dell’RNA ribosomiale durante la traduzione proteica.
Reticolo endoplasmatico
Il reticolo endoplasmatico (RE) è un complesso di membrane ripiegate, impilate e connesse tra loro, che occupa la maggior parte del citoplasma cellulare. Forma un compartimento chiuso delimitato dalla membrana e il suo spazio interno è chiamato lume.
Possiamo distinguere due tipi di RE:
- Il reticolo endoplasmatico rugoso (RER), che è ricoperto di ribosomi sulla sua faccia esterna, assumendo una forma sacculare. La maggior parte delle proteine passa attraverso le sue membrane e si accumula nelle cisterne. Inoltre, alcune reazioni di sintesi di carboidrati avvengono all’interno del lume del RER, dove enzimi catalizzano la glicosilazione, ovvero l’aggiunta di zuccheri alle proteine neoformate. Le vescicole si staccano dal RER per essere trasportate all’apparato del Golgi durante il processo di indirizzamento delle proteine. Il RER è coinvolto nella sintesi di proteine destinate alla secrezione (proteine che saranno rilasciate all’esterno della cellula, destinate ai lisosomi o alla membrana plasmatica).
- Il reticolo endoplasmatico liscio (REL), organizzato in una rete di tubuli senza ribosomi associati. Svolge un ruolo chiave nel metabolismo catabolico, tra cui la detossificazione delle sostanze nocive e la liberazione di glucosio dal glicogeno. Tuttavia, la sua funzione più importante è quella anabolica, essendo coinvolto nella sintesi di lipidi, come il colesterolo e altri steroidi.
Nelle cellule del muscolo scheletrico, il reticolo endoplasmatico liscio, noto come reticolo sarcoplasmatico, svolge una funzione specifica. In queste cellule, immagazzina ioni calcio che vengono rilasciati nel citosol in risposta a uno stimolo da parte di un motoneurone alla giunzione neuromuscolare, avviando così il processo di contrazione muscolare.
Apparato di Golgi
L’apparato di Golgi prende il nome dal medico italiano Camillo Golgi, che lo ha scoperto. È costituito da una serie di cisterne membranose appiattite, impilate una sopra l’altra, coinvolte nella glicosilazione, ovvero nel processo di aggiunta di residui glucidici per la produzione di glicolipidi e glicoproteine.
Inoltre, attraverso l’aggiunta di specifici residui chimici come oligosaccaridi, gruppi fosfato o altri complessi, l’organulo dirige le biomolecole verso la loro destinazione.
Il suo ruolo è essenziale nella formazione dei lisosomi, dove le idrolasi vengono indirizzate alla loro destinazione tramite l’aggiunta di gruppi fosfato. L’apparato di Golgi è coinvolto anche nella sintesi di glicoproteine destinate alla secrezione o presenti nella membrana cellulare, nella sintesi di glicosamminoglicani (GAG) e nella sintesi di lipoproteine e lipidi complessi.
Nonostante ci possano essere lievi variazioni a seconda delle cellule studiate, la struttura di questo organulo è generalmente uniforme.
Il Golgi è formato da una serie di cisterne, solitamente da quattro a sette, che si dispongono una di fronte all’altra. Una faccia del Golgi guarda verso il nucleo, mentre l’altra faccia guarda verso la membrana cellulare. Le vescicole di trasporto arrivano alla faccia rivolta verso il nucleo, mentre i vacuoli di condensazione, contenenti i prodotti dell’attività dell’organulo, originano dalla faccia rivolta verso la membrana cellulare. Le vescicole di trasporto sono piccole vescicole con un diametro di 80-100 nm che trasportano le biomolecole da elaborare al Golgi. I vacuoli di condensazione, invece, si originano dalla faccia trans del Golgi e si fondono tra loro per formare macrovescicole o granuli di secrezione (che possono essere costitutivi o regolati, come nel caso dei neurotrasmettitori) e vescicole lisosomiali (le proteine destinate al lisosoma sono marcate con una glicosilazione di mannosio-6-fosfato).

Lisosomi
I lisosomi sono organelli simili a sacchetti, circondati da una singola membrana. Svolgono un ruolo fondamentale come sistema digerente all’interno della cellula, responsabili della degradazione di molecole esterne, macromolecole internalizzate tramite fagocitosi e macromolecole endogene, come le parti invecchiate della cellula.
La digestione di molecole o particelle esterne alla cellula è nota come eterofagia, mentre la digestione di molecole intracellulari è chiamata autofagia.
La degradazione avviene grazie all’azione di enzimi idrolitici presenti nell’organulo, noti come “idrolasi acide“. Questi enzimi sono particolarmente attivi in un ambiente caratterizzato da un basso valore di pH. In media, il pH del lisosoma si aggira intorno a 5. Il basso pH viene creato e mantenuto grazie a pompe protoniche che trasportano ioni H+ dal citoplasma.
Gli enzimi sono in grado di degradare proteine, lipidi e carboidrati nei loro costituenti elementari, che possono poi essere riutilizzati in altri processi cellulari o espulsi. La distruzione di questi organuli comporta la morte cellulare.
Si distinguono due tipi principali di lisosomi: i lisosomi primari, che si formano dall’apparato del Golgi e contengono gli enzimi idrolitici, e i lisosomi secondari, che contengono il materiale da digerire.
Perossisomi
I perossisomi sono organelli rivestiti da membrana che contengono enzimi essenziali per la detossificazione della cellula e il metabolismo dell’alcool (etanolo). Tra gli enzimi di maggior rilevanza presente nel perossisoma, spicca la catalasi, responsabile della decomposizione dell’acqua ossigenata in ossigeno e acqua. Gli enzimi presenti all’interno del perossisoma catalizzano diverse reazioni che producono perossido di idrogeno, il quale viene neutralizzato all’interno del perossisoma stesso grazie all’azione della catalasi, rendendo il perossisoma un ambiente sicuro per la cellula. Inoltre, i perossisomi sono coinvolti in altre importanti funzioni cellulari, come la sintesi di lipidi e la degradazione degli acidi grassi a catena molto lunga.
La presenza dei perossisomi è particolarmente significativa in cellule che svolgono attività metaboliche intense, come il fegato, in cui l’elaborazione di sostanze tossiche, compreso l’alcool, è una priorità.

Mitocondrio
I mitocondri sono gli organelli cellulari responsabili della respirazione cellulare, un processo fondamentale per la produzione di energia sotto forma di ATP. Essi sono abbondanti in tutti i tessuti metabolicamente attivi. Strutturalmente, i mitocondri sono composti da una membrana esterna altamente permeabile alle molecole con una massa fino a 5.000 Dalton, una membrana interna che forma creste introflesse per aumentare la sua superficie e uno spazio intermembrana tra le due membrane.
La membrana interna dei mitocondri, grazie alle sue creste introflesse, ospita i complessi respiratori della catena di trasporto degli elettroni, i traslocatori di metaboliti e l’ATP sintetasi. La composizione della membrana mitocondriale è diversa dalle altre membrane cellulari: contiene una maggiore quantità di proteine, non contiene colesterolo e presenta cardiolipine, simili a quelle presenti nei batteri.
La membrana interna racchiude la matrice mitocondriale, che ha una consistenza gelatinosa dovuta alla sua elevata concentrazione di proteine idrosolubili (circa 500 mg/ml). La matrice contiene numerosi enzimi, ribosomi di dimensioni simili a quelli dei batteri (70S) e molecole di DNA mitocondriale circolare a doppio filamento, che codifica per alcuni tRNA, rRNA e proteine necessarie al funzionamento dei mitocondri.
Il DNA mitocondriale ha un’origine procariotica, e alcune caratteristiche dei mitocondri, come la presenza di DNA simile a quello dei procarioti e di ribosomi 70S, suggeriscono un’origine endosimbiotica. Tuttavia, alcune componenti dei mitocondri sono codificate dal DNA nucleare, sintetizzate nel citosol e poi assemblate nei mitocondri stessi. Il DNA mitocondriale viene trasmesso di generazione in generazione tramite la linea materna, poiché i mitocondri presenti nello zigote derivano principalmente dalla cellula uovo. Il DNA mitocondriale ha una struttura circolare e contiene solo 37 geni, di cui 13 codificano per le subunità proteiche dei complessi della catena respiratoria e 24 codificano per le molecole indispensabili alla sintesi dell’RNA. Ogni mitocondrio contiene da due a dieci copie del genoma mitocondriale.

La presenza della catena di trasporto degli elettroni nei mitocondri, con la sua capacità di produrre radicali liberi, insieme all’assenza di istoni e ai limitati meccanismi di riparazione, rende il DNA mitocondriale vulnerabile ai danni. In effetti, il tasso di mutazioni del DNA mitocondriale è circa dieci volte superiore rispetto al DNA nucleare.
Il numero e la distribuzione dei mitocondri possono variare: ad esempio, negli epatociti possono raggiungere il numero di 1.000-1.500, mentre sono presenti circa 30.000 mitocondri nell’ovocita. Nei globuli rossi, invece, i mitocondri sono assenti poiché queste cellule ottengono energia principalmente attraverso la glicolisi.